大青杨纤维素合酶(PuCesA7) cDNA的克隆及反义表达载体的构建 
2东北林业大学林学院, 哈尔滨,150040
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2011 年, 第 9 卷, 第 116 篇 doi: 10.5376/mpb.cn.2011.09.0116
收稿日期: 2011年10月08日 接受日期: 2011年10月20日 发表日期: 2011年11月17日
本研究提取大青杨叶片总 RNA,经 RT-PCR 扩增得到约1.3 kb 的 cDNA 片段,将其克隆到 pMD18-T 载体。经测序,该片段长1 304 bp,与欧洲颤杨 PtrCesA7 全长 cDNA 序列比对,核苷酸相似性为97%。克隆到载体上的 PuCesA7 片段用 SacⅠ和 KpnⅠ双酶切,反向插入到植物表达载体 pROKⅡ 的 P35S 启动子和 Tnos 终止子之间,成功构建 pROKⅡ-P7 SacⅠKpnⅠ反义表达载体。利用该植物表达载体对大青杨的遗传转化正在进行中,以确定 PuCesA7 基因的初步功能。
纤维素是自然界中含量最多,分布最广的一种多糖,从原始的单细胞生物如海藻、鞭毛和细菌等到高大的乔木到处可见。植物体内纤维素的含量因植物种类而差异显著,如棉花、木棉种子的绒毛中纤维素含量可达到95%~99%,是天然的最纯纤维素;麻、亚麻的韧皮纤维中含有的纤维素含量约为80%~90%;其次是木材和竹材,纤维素含量约为40%~50%。拟南芥基因组测序工作的完成推动了植物纤维素合成酶基因功能的深入研究。随着对拟南芥CesA基因家族了解的越来越多,针对有限的几种纤维素合酶基因已应用多种遗传学方法在其功能研究上取得了初步的成果,并提出了纤维素在生物体内合成的可能途径。第1个林木纤维素合酶基因是美国密西根大学的学者在2000年从欧洲颤杨中克隆获得的PtrCesA1基因(Wu et al., 2000),随后又发现PtrCesA2,PtrCesA3,PtrCesA4,PtrCesA5,PtrCesA6和PtrCesA7等基因(Samuga et al., 2002; Kalluri et al., 2003; 2004)。通过分析以上基因的表达时期和部位,认为PtrCesA1,PtrCesA2和PtrCesA3基因可能存在于同一个纤维素合酶复合体(CSC)中,与细胞次生壁的合成有关(Samuga et al., 2004);PtrCesA4,PtrCesA5和PtrCesA7基因可能与细胞初生壁的形成有关(Kalluri et al., 2004);PtrCesA6与茎的伸长生长有关(Liang et al., 2004)。Ranik和Myburg (2006)克隆获得6个巨桉纤维素合酶的全长cDNA。定量PCR结果显示:EgCesA1,EgCesA2和EgCesA3在木质部等次生生长组织中的转录水平较高,而在叶片等初生生长的组织中较低;EgCesA4和EgCesA5在初生壁快速分化组织中表达上升;EgCesA6在所有的组织中表达均较弱(Ranik and Myburg, 2006)。但有关纤维素合酶的具体合成机制尚有许多问题有待深入研究。
大青杨是我国东北地区森林环境下重要的杨树乡土树种,具有适应能力强、生长快、干型好、木材轻软、材质白色、致密、耐朽力强等优点,是造纸及胶合板材极好的原料。作为黑龙江省东部山区速生丰产林的主栽品种之一,生长量最高,人工栽培技术掌握最充分,在山地森林环境下造林(更新)成效最明显,特别是在幼林阶段,是很有希望的短轮伐期工业用材树种。了解纤维素合酶的生物合成机制,希望利用基因工程等手段来增加纤维素含量以改善大青杨的品质,培育出更适于造纸的新型树种。
1结果与分析
1.1 cDNA片段的获得
以反转录获得的第一条链cDNA为模板进行扩增,结果可见长度为1 304 bp的片段(图1)。用NCBI中的BLAST程序将获得的cDNA序列在GenBank中进行序列同源性比对分析,结果显示与欧洲颤杨PtrCesA7基因全长cDNA序列序列相似性为97%,将此序列命名为PuCesA7 (GenBank登录号HQ288419)。
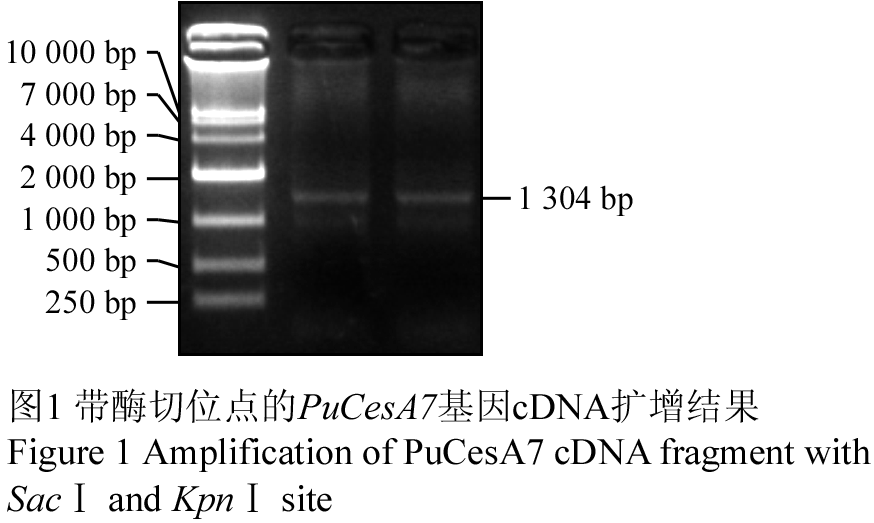 图1 带酶切位点的PuCesA7基因cDNA扩增结果 Figure 1 Amplification of PuCesA7 cDNA fragment with SacⅠ and KpnⅠ site |
1.2 T-载体克隆及鉴定
重组T-载体经PCR初步鉴定后,选取阳性克隆进行双酶切鉴定(图2)及序列测定。结果表明,酶切片段为目标基因。将经双酶切的PuCesA7 cDNA片段命名为P7SacⅠKpnⅠ,重组T-载体命名为pMD18-T-P7SacⅠKpnⅠ。
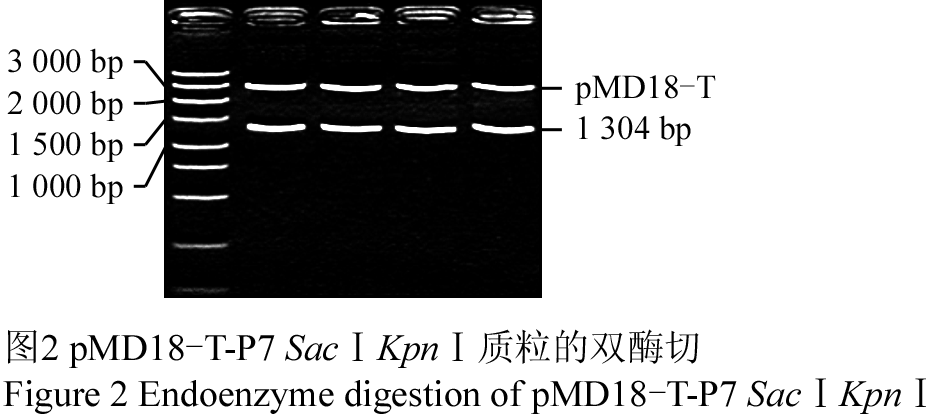 图2 pMD18-T-P7 SacⅠKpnⅠ质粒的双酶切 Figure 2 Endoenzyme digestion of pMD18-T-P7 SacⅠKpnⅠ |
1.3反义表达载体构建
对重组质粒pMD18-T-P7SacⅠKpnⅠ和植物表达载体pROKⅡ进行双酶切(SacⅠ, KpnⅠ),将目的片段反向连接到表达载体上。对构建的反义表达载体进行PCR和双酶切验证,琼脂糖凝胶电泳结果显示可切出目的片段(图3),测序结果亦表明目的片段与植物表达载体连接正确。反义表达载体构建策略见(图4),将构建的反义表达载体命名为pROKⅡ-P7SacⅠKpnⅠ。
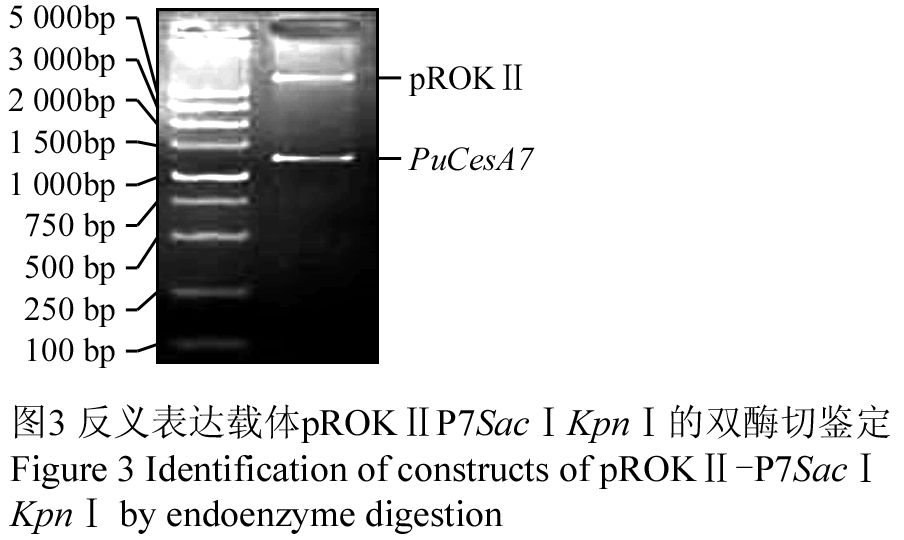 图3 反义表达载体pROKⅡP7SacⅠKpnⅠ的双酶切鉴定 Figure 3 Identification of constructs of pROKⅡ-P7SacⅠKpnⅠ by endoenzyme digestion |
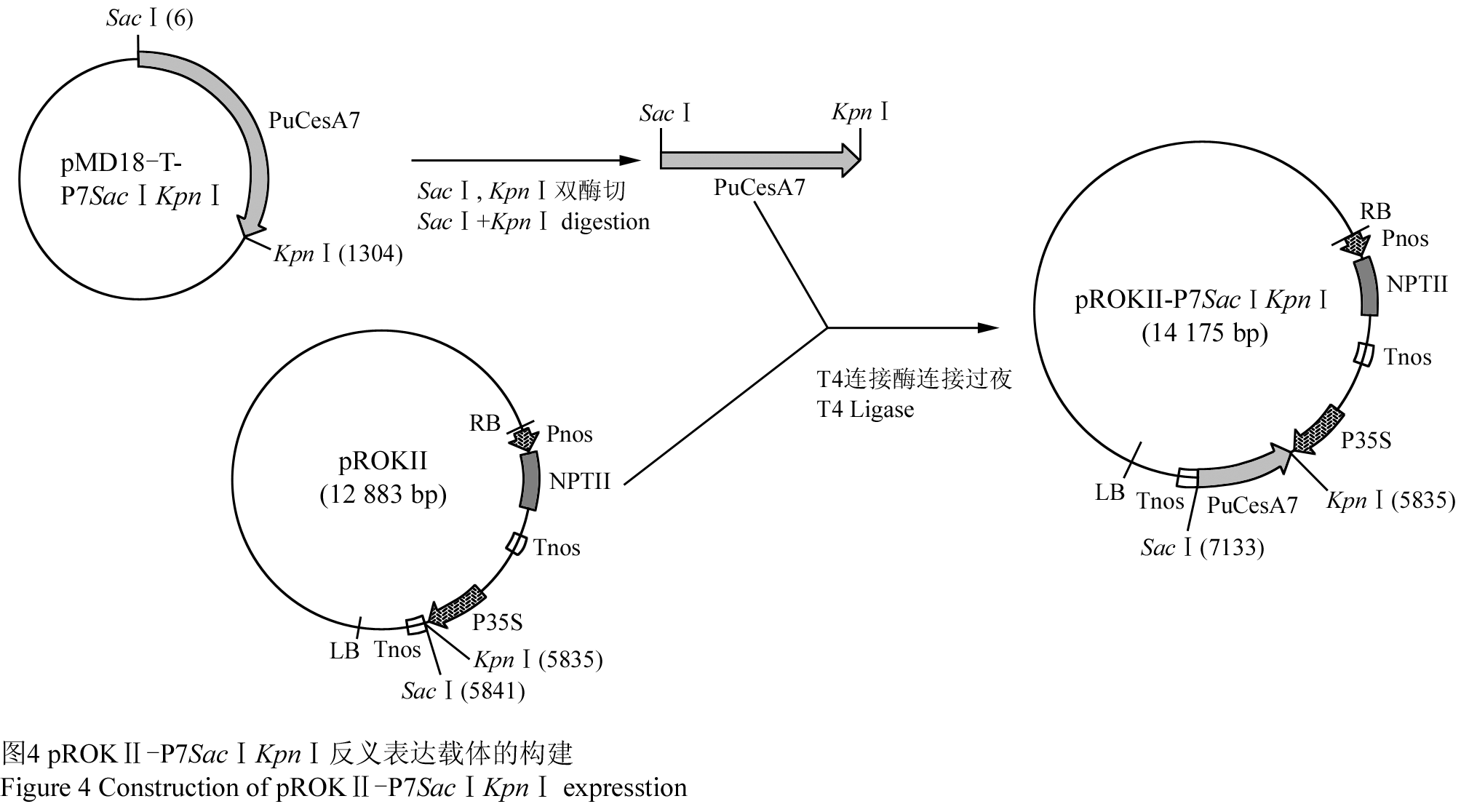 图4 pROKⅡ-P7SacⅠKpnⅠ反义表达载体的构建 Figure 4 Construction of pROKⅡ-P7SacⅠKpnⅠ expresstion vector |
2讨论
反义RNA的设计没有必要使用全长的cDNA序列,也不需要了解目的基因的详细调控知识(孟博和朱大海, 2001, 国外医学遗传学分册, 24(6): 289-292)。对真核生物而言,针对5'非编码区设计的反义RNA要优于针对编码区的设计,可能是因为这些非编码序列能够调控编码序列RNA和蛋白质的合成以及蛋白质空间结构的形成。由于反义 RNA可专一抑制特定基因的表达而不影响其它基因,因此只要将一个无启动子的克隆序列以反向形式与某启动子连接,并通过载体将之导入受体细胞中,就可使细胞合成特异性的反义RNA,进而调控相关基因的生物功能(许本波等, 2003),常用来研究基因失活与表型变化间的相互关系。
生物体内纤维素的合成都与β-1,4糖苷键有关,这是一个相当复杂的过程,并受到多种因素的调控。不同植物的纤维素在合成过程中,需要多种纤维素合酶基因的共同调控,并且这些纤维素合酶基因的作用也是不同的。Burton等(2000)利用病毒诱导植物纤维素合成酶基因沉默的方法将3个与CesA基因相关的cDNA片段分别插入马铃薯X病毒载体中,接种到烟草(Nicotiana benthamiana)来研究CesA基因的功能,试验结果出现受感染的植株节间缩短、小叶和侏儒的症状,并且叶表面由于缺少纤维素而表现出细胞膨胀,叶片中的多聚糖含量降低了25%。这些现象说明cDNA片段沉默了一个或多个纤维素合成酶基因的表达,导致CesA基因转录水平的降低(Burton et al., 2000)。Burn等(2002)通过构建反义表达载体并转化拟南芥进行纤维素合成酶基因的功能分析,结果表明拟南芥CesA1,CesA2,CesA3 3个基因存在连锁表达的情况,由此推断这3个基因可能组合在一个纤维素合酶复合体(CSC)中并参与纤维素的生物合成,主要功能与细胞初生壁的合成相关(Burn et al., 2002)。本研究成功构建了大青杨纤维素合酶PuCesA7基因的反义表达载体,下一步工作将通过转基因大青杨的表型差异来确定纤维素合酶PuCesA7基因的初步功能以及与其它纤维素合酶基因的相互关系。
3材料与方法
3.1材料
取大青杨幼嫩叶片,采用改良2× CTAB法提取叶片总RNA (许雷等, 2010)。根据欧洲颤杨PtrCesA7基因全长cDNA序列设计一对带特异酶切位点的引物P7F (5'-GAGCTCGGATGAGGGCAGGCAGCCACTTTC-3')和P7R (5'-GGTACCATGCTTCCCTGTTCTTTGACTTCT-3')。cDNA第一链的合成,采用20 μL反转录体系:5× Buffer (4 μL),RNase Inhibitor (1 μL),Dntp (2.5 mmol/Leach) (4 μL),Oligo dT18 (1 μL),Random Primer (1 μL),AMV反转录酶XL (2 μL),总RNA (2 μL),RNase-free H2O (5 μL)。54℃延伸至少2 h,冰上放置终止反应。T4 DNA连接酶,限制性内切酶(SacⅠ, KpnⅠ),AMV反转录酶XL,pMD18-T载体,LA Taq聚合酶购自TAKARA公司;胶回收试剂盒,质粒提取试剂盒为PROMEG公司。
3.2 cDNA片段的扩增
建立20 μL PCR反应体系:10× Buffer (2 μL),dNTP (2.5 mmol/L each) (1 μL),5'特异引物(10 μmol/L) (0.5 μL),3'特异引物(10 μmol/L) (0.5 μL),cDNA第一条链(1 μL),LA Taq聚合酶(0.3 μL),ddH2O (14.7 μL)。扩增程序为94℃下预变性5 min;94℃下变性30 s,57℃退火45 s,72℃延伸4 min,35个循环;72℃延伸15 min。对PCR产物进行1.2%的琼脂糖凝胶电泳,回收目的片段。
3.3 T-载体克隆鉴定
目的片段与T-载体连接按试剂盒指导操作,重组质粒转化至E. coli DH5α感受态细胞。经过蓝白斑筛选、特异引物PCR鉴定筛选出阳性克隆;初步鉴定为阳性克隆的质粒进行双酶切(SacⅠ, KpnⅠ),能切出相应大小片段的克隆即为阳性克隆;对阳性克隆作DNA序列测定,测序工作由北京六合华大基因科技股份有限公司测序。
3.4反义表达载体的构建和鉴定
选择植物表达载体pROKⅡ质粒,该载体的选择标记为NPTⅡ,即新霉素磷酸转移酶(Neomycin Phosphotransferase Ⅱ)。分别对重组质粒和植物表达载体pROKⅡ进行双酶切(SacⅠ, KpnⅠ),回收目的片段。用T4 DNA ligase连接目标片段与载体,将连接产物转化E. coli DH5α感受态细胞,提取质粒。经PCR初步鉴定后,再用SacⅠ,KpnⅠ双酶切鉴定阳性克隆。阳性克隆送测序鉴定。
作者贡献
许雷为本研究的构思者及负责人,试验设计和试验研究的执行人;方连玉参与试验设计,试验结果分析,数据分析及论文初稿的写作;刘一星指导试验设计,数据分析,论文写作与修改工作。全体作者都阅读并同意最终的文本。
致谢
本研究由中央高校基本科研业务费专项资金(41401005)及林业工程学科创新引智基地项目(B080160)资助。
参考文献
Burn J.E., Hocart C.H., Birch R.J., Cork A.C., and Williamson R.E., 2002, Functional analysis of the cellulose synthase genes CesA1, CesA2, and CesA3 in Arabidopsis, Plant Physiol., 129(2): 797-807
Burton R.A., Gibeaut D.M., Bacic A., Findlay K., Roberts K., Hamilton A., Baulcombe D.C., and Fincher G.B., 2000, Virus-induced silencing of a plant cellulose synthase gene, Plant Cell, 12(5): 691-706
Kalluri U.C., and Joshi C.P., 2003, Isolation and characterization of a new, full-length cellulose synthase cDNA, PtrCesA5 from developing xylem of aspen trees, J. Exp. Bot., 54(390): 2187-2188
Kalluri U.C., and Joshi C.P., 2004, Differential expression patterns of two cellulose synthase genes are associated with primary and secondary cell wall development in aspen trees, Planta, 220(1): 47-55
Liang X., and Joshi C.P., 2004, Molecular cloning of ten distinct hypervariable regions from the cellulose synthase gene superfamily in aspen trees, Tree Physiol., 24(5): 543-550
Ranik M., and Myburg A.A., 2006, Six new cellulose synthase genes from Eucalyptus are associated with primary and secondary cell wall biosynthesis, Tree Physiol., 26(5): 545-556
Samuga A., and Joshi C.P., 2002, A new cellulose synthase gene (PtrCesA2) from aspen xylem is orthologous toArabidopsis AtCesA7(irx3) gene associated with secondary cell wall synthesis, Gene, 296(1-2):37-44
Samuga A., and Joshi C.P., 2004, Cloning and characterization of cellulose synthase-like gene PtrCSLD2 from developing xylem of aspen trees, Physiol. Plant., 120(4): 631-641
Wu L., Joshi C.P., and Chiang V.L., 2000, Axylem-specific cellulose synthase gene from aspen (Populus tremuloides) is responsive to mechanical stress, Plant J., 22(6): 495-502
Xu B.B., Zhang X.K., and Li J.N., 2003, Application of antisense RNA technique in crop genetic improvement, Zhongguo Nongxue Tongbao (Chinese Agricultural Science Bulletin), 19(3): 84-88 (许本波, 张学昆, 李加纳, 2003, 反义RNA技术在现代作物遗传改良中的应用, 中国农学通报, 19(3): 84-88)
Xu L., Liu Y.X., and Fang L.Y., 2010, Method of Extracting RNA Quickly from Leaves of Populus ussuriensis kom., Shengwu Jishu Tongxun (Letters in Biotechnology), 21(6): 857-860 (许雷, 刘一星, 方连玉, 2010, 大青杨叶片总RNA的快速提取方法, 生物技术通讯, 21(6): 857-860)